
Friedrich Engels believed that labor created humans, but modern scientists argue that love, children's curiosity, and long-distance running played a much more significant role in the formation and development of our species. In his lecture series for Qalam, paleontologist Alexander Markov explains how and why Homo sapiens emerged.
For millennia, philosophers, priests, and poets have debated whether the spirit or body has primacy in humans. These debates are vibrant and fascinating, but they rarely lead to conclusions. However, when biologists joined the discussion, answers slowly began to emerge, and the past started to solidify from out of the mists of time. This course is dedicated to human evolution and our environment’s role in it.
Among those who are unfamiliar with evolutionary biology, one common misconception is that evolutionary processes can be understood as an organism’s reaction to changes in the external environment, following the logic of ‘It got cold, and thick fur grew’. However, such a view is one-sided and incomplete. Understanding biological evolution as a simple reaction to environmental challenges is incorrect. Of course, the environment definitely influences evolution, which is hard to dispute. Living organisms adapt to environmental conditions through biological (genetic) evolution, and humans do this through cultural evolution as well (following the logic of ‘It got cold, so let's wrap ourselves in furs’). However, the structure of organisms does not arise directly from the conditions in the environment they are adapting to. After all, there are countless ways to adapt to the same environment.
For example, one could say that a bird’s wings, feathers, and flight are a ‘consequence’ of the fact that the Earth's atmosphere has a specific density, making it possible to fly in it the way birds do. However, such a statement would be of little value because there are many different ways to adapt to life in this atmosphere and to move through the air. Spiders fly on silk threads; birds flap their feathered wings; and bats, pterosaurs, and dragonflies have wings of entirely different designs, while humans invented airplanes.
CULTURAL HUMANS
The most significant events in human evolution occurred during the Pleistocene epoch or the Ice Age, which occurred from 2.58 million to 11,700 years ago, when periodic glaciations began in the Northern Hemisphere, and the climate in Africa, where our ancestors lived, became more arid, with the forested areas shrinking and savannas expanding. Many features of our ancestors' evolution were related to their emergence from the forests and to their adaptation to life on the savanna.
However, this does not mean that the Ice Age caused the emergence of humans as is sometimes suggested in lower-quality popular science books.
The Ice Age affected all animals equally. In Africa, numerous species adapted in completely different ways to the reduction of forests and the growth of savannas. The ancestors of baboons, for instance, also transitioned from forests to savannas and adapted but did not become intelligent beings.
In addition to biological evolution, which is based on genetic inheritance, humans also have cultural evolution based on social learning, the non-genetic transmission of knowledge, skills, customs, beliefs, and technologies from person to person. Our cultural adaptations (such as building shelters, wearing clothes, cooking food over a fire, et cetera) radically changed the very ‘external environment’ to which our bodies adapted through biological evolution. Culture set the direction and strength of natural selection, which shaped our bodies, and changes in our bodies (especially in such a crucial part as the brain) influenced the further development of culture.

Figures of Stone Age hominids making tools at the National Museum of Mongolian History/Wikimedia Commons
Taking this context into account, we will discuss the current data on the development of important distinguishing features of humans—including bipedalism, strong long-distance running abilities, lack of fur, an abundance of sweat glands, as well as the tendency to form stable mating pairs and a prolonged childhood—within the framework of this course. Additionally, we will try to understand the reasons for the extraordinarily rapid (by evolutionary standards) increase in brain size in our ancestors.
WHY WALK UPRIGHT AT ALL?
The story of human evolution often begins with bipedalism, quite literally, walking on two legs. Among all the apes, only humans use this as their primary method of movement, and it is one of our most unique characteristics. Interestingly, bipedalism evolved before all other traits that differentiate us from chimpanzees (our closest living relatives).
It’s true that chimpanzees, like gorillas (our slightly more distant relatives), sometimes stand upright and walk on two legs. However, they do this only occasionally, around 5–10 per cent of the time. For all apes, except humans, this posture is uncomfortable and mostly unnecessary: their arms are so long that, with a slight hunch, they are already on all fours. It’s easier for them to walk by leaning on their knuckles, fists, or palms.
Today, there is only one species of bipedal ape on Earth—Homo sapiens, that is, us.
But in the past, there were many more. Paleontologists have determined that a particular group of bipedal apes emerged around 7 million years ago in Africa. They thrived for a long time and were quite diverse, consisting of several genera and around fifteen to twenty species. Initially, their brains were no larger than those of chimpanzees, and they likely did not surpass chimpanzees in intellectual abilities.
Unfortunately, all these bipedal apes with chimpanzee-sized brains eventually became extinct, except for those which evolved into humans. They all belonged to the ‘human’ evolutionary lineage, meaning they were closer relatives of humans than chimpanzees. We will refer to the representatives of this lineage as hominids (In Latin ‘hominidae’ meaning ‘human-like’; the singular of this word is ‘hominid’). Hominids first appeared in Africa, and their entire early evolution occurred there. Darwin first proposed the idea that the fossil ancestors of humans lived on the African continent in his book The Descent of Man, and Selection in Relation to Sex. At the time, when scientists had not yet found a single bone of anything remotely resembling a ‘missing link’ between apes and humans, Darwin’s hypothesis seemed incredibly bold.

Charles Darwin's 1837 sketch. His first diagram of an evolutionary tree from his First Notebook on Transmutation of Species /Wikimedia Commons
Hominids are characterized by a key common trait—walking on two legs. There are at least as many hypotheses explaining the transition to bipedalism as there are reasons why apes sometimes stand upright. Apes stand on two feet when crossing shallow water bodies. Could our ancestors have become bipedal because they spent a lot of time in the water? Indeed, there is such a hypothesis. Male apes stand up straight and display their penises when courting females. Could our ancestors have wanted to show off their genitals all the time? There's a hypothesis for that, too! Female apes sometimes walk on two legs while holding a baby against their belly (if the baby isn’t clinging to their mother’s back, grabbing her fur). Maybe our ancestors needed to carry two babies at once and freed their hands for that purpose? Well, there’s a hypothesis for that as well …

Bonobo chimpanzee (Pan paniscus) carrying her young in a bipedal position. Zaire, Congo/Getty Images
But that’s not all. Scientists have also suggested that our ancestors wanted to increase their field of vision (which became especially important after leaving the forest for the savanna). Or perhaps they wanted to reduce the body surface exposed to the sun, again, after transitioning to the savanna. How do we choose the correct idea from all these options? Or could several ideas be right at the same time? It's hard to say. Entire articles and even books have been devoted to arguments in favor of each of these hypotheses, but none have direct evidence.
In such cases, I believe it is best to favor hypotheses that have additional explanatory power—those that not only explain bipedalism but also other unique characteristics of hominids. This way, we need to rely on far fewer debatable assumptions. We will discuss one such hypothesis, but first, let’s become better acquainted with the facts.
THE EARLIEST BIPEDS
It was traditionally believed that the last common ancestor of humans and chimpanzees preferred to walk on all fours, much like chimpanzees do today. It was thought that this original (primitive) method of locomotion was retained by chimpanzees, gorillas, and orangutans. At the same time, in our evolutionary line, it was replaced by bipedalism as a result of leaving the forest for the open savanna. However, suspicions later arose that perhaps the last common ancestor of humans and chimpanzees, if not fully bipedal, at least showed more inclination toward upright walking than modern chimpanzees and gorillas. This possibility is hinted at by new paleoanthropological discoveries.

Western lowland gorilla juvenile female 'Bokata' aged 6 years walking bi-pedally to cross a river. Bai Hokou, Dzanga Sangha Special Dense Forest Reserve, Central African Republic/Getty Images
Over the last thirty years, archeologists have discovered the bones of several species of very ancient hominids in Africa, dating from 7 to 4 million years ago. The classification of these forms remains debated. Although they have been described as members of three new genera (Sahelanthropus, Orrorin, Ardipithecus), some experts believe that certain species should be grouped together or with the later genus Australopithecus.i from the Latin and Greek ‘southern ape’, a genus of primates that lived from about 4.2 to 1.8 million years ago
from the Latin and Greek ‘southern ape’, a genus of primates that lived from about 4.2 to 1.8 million years ago

22 July 2022, Dusseldorf, Germany: Lucy - Australopithecus afarensis human ancestor by Charles Darwin Evolution Theory and anthropology science/Alamy
What’s most intriguing about these earliest hominids is that they all seem to have walked on two legs, though not as confidently as we do. This is indicated by a forward-shifted foramen magnum (the large opening at the base of the skull), meaning the spine was attached to the skull from below rather than from the back, which clearly points to bipedal locomotion. Therefore, our ancestors were walking upright a very long time ago. Almost immediately after the human and chimpanzee lines diverged (around 7 million years ago), members of ‘our’ line were already walking on two legs. Or perhaps that happened even earlier? What if the common ancestors of humans and chimpanzees already preferred walking on their hind limbs, and the current knuckle-walking style of chimpanzees developed later? They lived not in open savannas but in sparsely forested areas or mixed landscapes where forest patches alternated with open spaces. This doesn’t necessarily contradict the old theory that bipedalism developed as forest dwellers transitioned to open areas. The only difference, according to new data, is that this transition was gradual and not abrupt.
OUR ORANGUTAN COUSINS
Recent evidence suggests that bipedal walking might be rooted in the locomotion patterns of apes that developed while they were living in trees. For example, it was recently demonstrated that the way orangutans move on two legs while holding onto branches with their hands most closely resembles human walking.
An earlier idea suggested that the skeleton and musculature of our ancestors were predisposed to bipedal walking due to their tree-climbing abilities. In this position, the body is vertically aligned, and the legs make movements similar to walking. However, anthropologist Robin Crompton from the University of Liverpool, along with his colleagues Susanne Thorpe and Roger Holder from the University of Birmingham, believe that it’s difficult to move to human bipedal walking from vertical tree-climbing or the gait of chimpanzees and gorillas. The mechanics of these movements differ significantly. For instance, the knees of chimpanzees and gorillas almost never fully straighten. As we know, these apes sometimes walk on two legs, but they keep their legs bent. There are also other differences between their gait and that of humans. Orangutans, on the other hand, are the most arboreal of the great apes, and Crompton and his colleagues have studied their behavior in the forests of Sumatra.
The anthropologists recorded 2,811 individual instances of orangutan movement in the treetops. For each case, they noted the number of supports (branches), their thickness, and the method of movement. Orangutans use three main modes: bipedal walking (holding onto something with their hands), quadrupedal movement (grasping branches with both hands and feet), and suspension (hanging from branches with their arms while occasionally grabbing with their feet).
It was found that the method of movement depends on the number and thickness of the supports. On single, thick, sturdy branches, orangutans usually move on all fours; on medium-diameter branches, they move with their arms. On thin branches, they prefer to carefully walk on their feet, using their hands to hold onto an additional support. In this case, their gait is very similar to that of humans, and their knees fully straighten. This method of movement appears to be the safest and most efficient when navigating thin, flexible, and unreliable branches. An additional advantage is that one hand remains free to pick fruit.

Luna, an orangutan, walks on a rope, while her nearly 3-month-old baby, Stella, holds on to her at Busch Gardens. Tampa, Florida, USA/Jefferee Woo/Tampa Bay Times via ZUMA Press Wire/Alamy
The ability to walk on thin branches is no small feat for arboreal apes. Thanks to this skill, they can move freely across the forest canopy and transition from tree to tree without descending to the ground. This significantly conserves energy, reducing the effort needed to forage. Therefore, such an ability would be strongly favored by natural selection.
The ancestors of orangutans diverged from our ancestors earlier than gorillas, and gorillas diverged earlier than chimpanzees. Researchers suggest that walking on two legs across thin branches was originally inherent to the distant ancestors of all great apes. Orangutans, living in the humid tropical forests of Southeast Asia, retained and further developed this skill, while gorillas and chimpanzees lost it, instead evolving knuckle-walking on all fours and occasionally using a semi-bent bipedal gait. This shift could have been influenced by the periodic drying out of the tropical forests in Africa and the spread of the savannas. The human evolutionary line learned to walk on the ground in the same way they moved on thin branches, straightening their knees.

Evolution/Wikimedia Commons
According to Crompton and his colleagues, their hypothesis explains the two groups of facts that seem puzzling from the perspective of other bipedalism origin theories. First, it explains why species closely related to the common ancestor of humans and chimpanzees (like Sahelanthropus, Orrorin, and Ardipithecus) already exhibit the distinct skeletal traits of bipedalism despite having lived in the forest rather than the savanna. Second, it resolves the apparent contradiction in the limb structure of australopithecines, the bipedal apes from which the first humans (Homo) emerged. Australopithecines had legs well-adapted for bipedal walking, yet their hands were long, strong, and better suited for life in trees and grasping branches.

Australopithecus afarensis model reconstruction at the Smithsonian National Museum of Natural History/Wikimedia Commons
Thus, it is possible that both humans and orangutans retained the ancient bipedal gait of their distant ancestors, while gorillas and chimpanzees lost it and developed something new—knuckle walking. In this sense, humans and orangutans could be considered ‘primitive’ in terms of their gait, while chimpanzees and gorillas are ‘evolutionarily advanced’.
WHAT BEAUTIFUL ARDI REVEALED TO US
Still further clarity on the origins of bipedalism comes from Ardi, the most ancient and well-studied hominid to date.
In October 2009, a special issue of the Science journal dedicated to the comprehensive study of Ardipithecus (a bipedal ape that lived in northeastern Ethiopia 4.4 million years ago) bones was published. And based on several teeth and jaw fragments, the species Ardipithecus ramidus was first described in 1994. In the following years, the collection of Ardipithecus bones was significantly expanded, and the most remarkable discovery was a substantial portion of the skeleton of a female individual, which scientists proudly introduced to the public and media under the name ‘Ardi’. Officially, Ardi is cataloged as ‘Skeleton ARA-VP-6/500’.
4.4 million years is the age of the oldest studied representative of the hominid family.
The special issue of Science featured eleven articles summarizing years of work by a large international research team. The publication of these articles and their central figure, Ardi, were widely publicized. But this wasn't just hype—the study of Ardipithecus bones truly allowed for a more detailed and accurate reconstruction of the early stages of hominid evolution.
It was confirmed that Ardipithecus ramidus is an excellent candidate for the role of a transitional link between the common ancestor of humans and chimpanzees (to which Orrorin and Sahelanthropus were also likely closely related) and later hominids, the australopithecines, from whom the first members of the human genus (Homo) emerged.
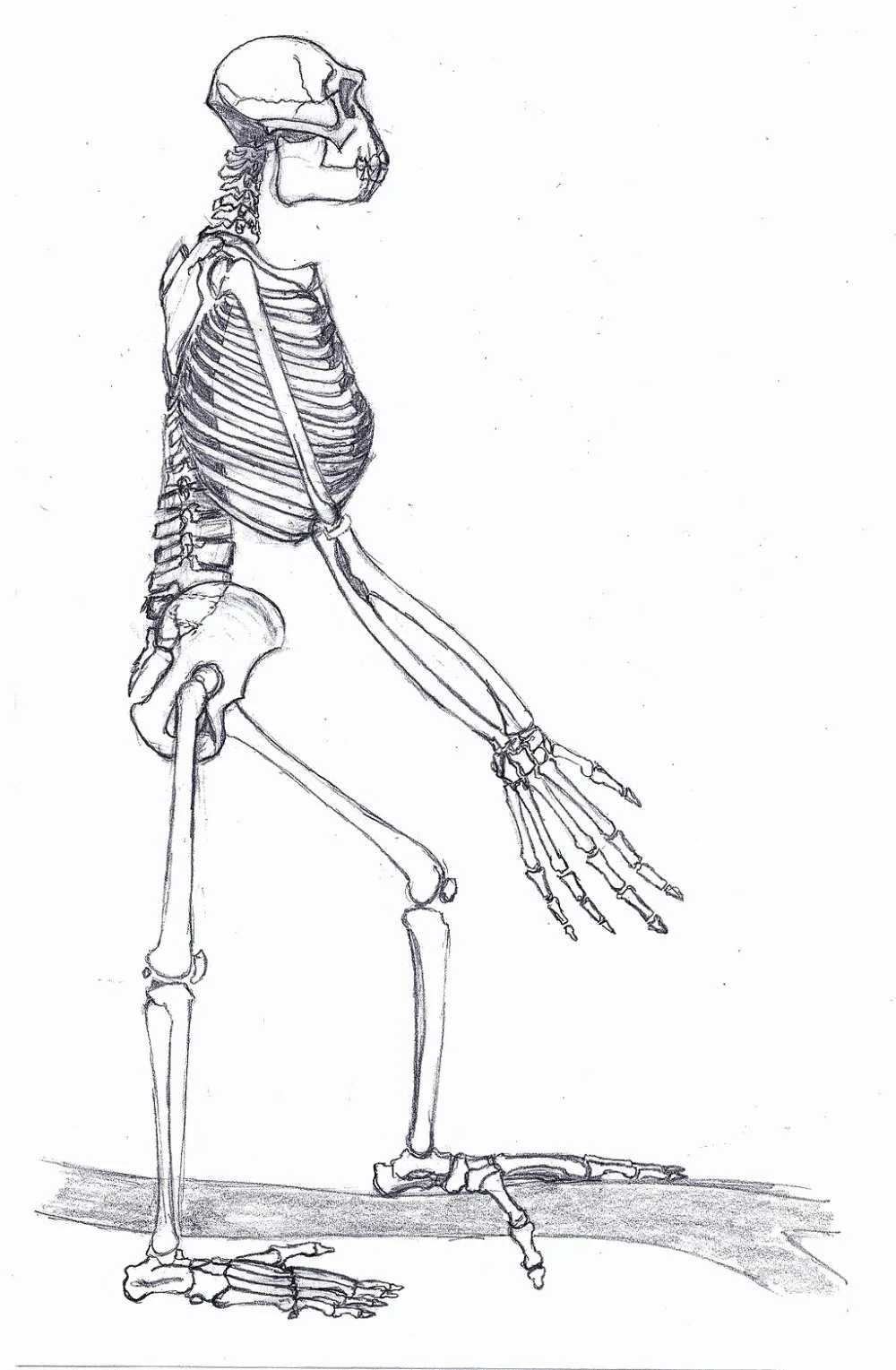
Ardipithecus Gesamt/Wikimedia Commons
Until 2009, the most ancient, well-studied hominid was Lucy, an Australopithecus afarensis who lived about 3.2 million years ago. All older species (in ascending order of age: Australopithecus anamensis, Ardipithecus ramidus, Ardipithecus kadabba, Orrorin tugenensis, Sahelanthropus tchadensis) were studied from fragmentary material. Consequently, our understanding of their structure, lifestyle, and evolution remained incomplete and imprecise. However, in 2009, the honorable title of ‘the most ancient well-studied hominid’ was ceremoniously passed from Lucy to Ardi.
The bones of the Ardipithecus ramidus were found in a single layer of sediment about 3 meters thick enclosed between two volcanic layers. The age of these layers was determined using the argon-argon method and was the same (within the margin of error): 4.4 million years. This means that the bone-bearing layer formed (as a result of floods) relatively quickly—in a maximum of 100,000 years, but most likely over a few thousand or even hundreds of years.

Ardipithecus ramidus genus of extinct hominin. Fossilized partial female hominid skeleton. Extinct primate Found in Awash Ethiopia/Alamy
Fragments of Ardi's skeleton were scattered over an area of about 3 square meters. The bones were exceptionally fragile, and extracting them from the rock required considerable effort. The cause of Ardi's death has not been determined. She was not eaten by predators, but it seems her remains were thoroughly trampled by large herbivores. The skull, in particular, was crushed into many fragments.
The remains of various animals and plants were found along with the bones of Ardipithecus ramidus. The plant remains were predominantly from forest species, and the animals were mostly those that fed on tree leaves or fruits (not grass). Based on these findings, it is thought that Ardipithecus ramidus lived not in the savanna but in a wooded area where dense forest patches alternated with more open spaces. The ratio of carbon isotopes 12C and 13C in the tooth enamel of five Ardipithecus ramidus individuals indicates that they primarily ate forest resources, not savanna plants (savanna grasses have a higher 13C isotope content). This differentiates Ardipithecus from its descendants, the australopithecines, who obtained 30 to 80 per cent of their carbon from open-space ecosystems (Ardipithecus got 10 to 25 per cent). However, Ardipithecus ramidus was not purely a forest dweller like chimpanzees, whose food is nearly 100 per cent forest-derived.
The fact that Ardipithecus ramidus lived in the forest seems, at first glance, to contradict the old hypothesis that early hominid evolution and the development of bipedalism were linked to moving out of the forest into the savanna. Similar conclusions had been made earlier in the study of Orrorin and Sahelanthropus, who also apparently walked on two legs but lived in forested areas. However, this situation can be viewed differently if we remember that the forests where early hominids lived were not very dense, and their bipedalism was not very developed. In other words, hominids transitioned from dense forests to open spaces gradually, and their walking ability improved gradually as well.
Ardi’s skull is similar to that of Sahelanthropus—both species are characterized by a small brain volume (300–350 cubic centimeters), a forward-shifted foramen magnum (indicating that the spine was attached to the skull from below rather than from behind as in humans), and less developed molars and premolars compared to chimpanzees and gorillas. It appears that the pronounced prognathism (the forward projection of the jaws) seen in modern African great apes is not a primitive trait and developed after their ancestors diverged from those of humans.
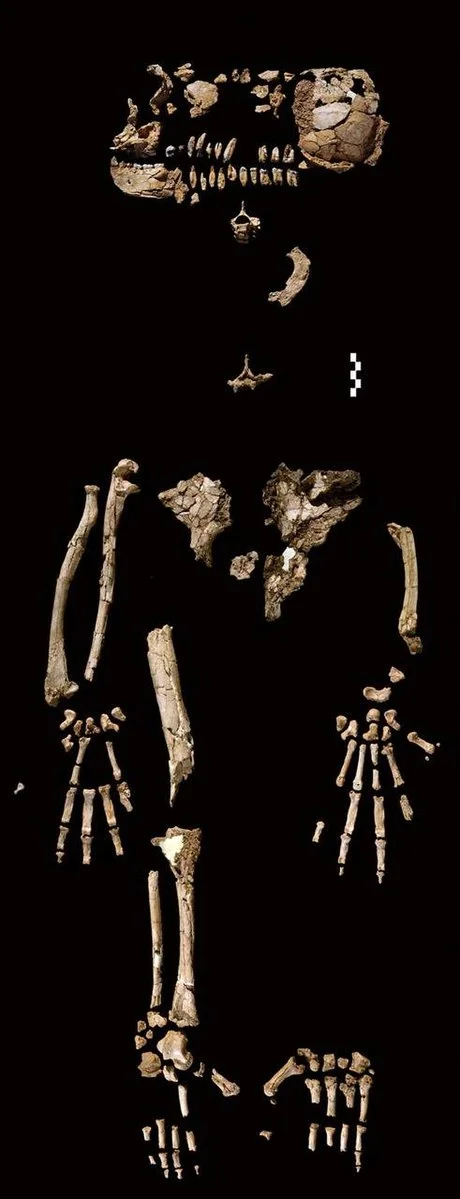
Skeletal fragments of Ardipithecis Ramidus/Wikimedia Commons
The teeth of Ardipithecus ramidus indicate that it was an omnivorous creature. The combination of features (the size of the teeth, their shape, enamel thickness, the nature of microscopic scratches on the tooth surface, isotopic composition) suggests that Ardipithecus ramidus did not specialize in any one diet, such as fruits, like chimpanzees do. It seems that Ardipithecus ramidus fed both in trees and on the ground, and its food was not overly tough.
One of the most important facts is that in male Ardipithecus ramidus, unlike in modern great apes (except humans), the canines were not larger than those of the females. Male apes actively use their canines both to intimidate rivals and as weapons. In the earliest hominids (Ardipithecus kadabba, Orrorin, Sahelanthropus), the males’ canines may also have been no larger than those of females, although the data is not sufficient for definitive conclusions. It is evident that in the human evolutionary line, sexual dimorphism in canine size disappeared very early. One could say that the males’ canines became ‘feminized’. In chimpanzees and gorillas, sexual dimorphism in canines reemerged, with males developing very large canines. In male bonobos, the canines are smaller than in other modern great apes, and bonobos are also characterized by the lowest levels of intraspecific aggression. Many anthropologists believe there is a direct link between canine size in males and intraspecific aggression. In other words, it can be hypothesized that the reduction of canines in our distant ancestors was linked to certain changes in social structure, such as a reduction in conflicts between males.
Ardi was about 120 cm tall and weighed about 50 kg
Ardi was approximately 120 centimeters tall and weighed about 50 kilograms. Male and female Ardipithecus individuals showed almost no size difference. This weak sexual dimorphism is also characteristic of modern chimpanzees and bonobos, with their relatively egalitarian relationships between the sexes. In contrast, gorillas exhibit strong sexual dimorphism, typically associated with polygamy and a harem system.
Ardi walked on two legs when on the ground, although less confidently than Lucy and her relatives, the australopithecines. At the same time, Ardi retained many specific adaptations for efficient tree climbing. Accordingly, the structure of Ardi’s pelvis and legs shows a combination of primitive (tree-climbing) and advanced (walking) features.
Ardi’s hands were exceptionally well-preserved (unlike Lucy’s hands), allowing for important conclusions. As we already know, it was long believed that the ancestors of humans, like chimpanzees and gorillas, walked using their knuckles. This unique form of locomotion is typical only of African great apes and orangutans; other monkeys usually walk on their palms. However, Ardi’s hands lacked the specific traits associated with knuckle-walking. Ardipithecus had hands that were more flexible and mobile than those of chimpanzees and gorillas and, in some respects, similar to those of humans. It is now clear that these traits are ‘primitive’, ancestral to hominids (and possibly to the common ancestor of humans and chimpanzees). The hand structure seen in chimpanzees and gorillas (which, incidentally, does not allow them to manipulate objects as skillfully as we do) is instead ‘advanced’ and specialized. The strong, grasping hands of chimpanzees and gorillas allow these large animals to move efficiently through trees but are poorly suited for delicate manipulation. Ardi’s hands allowed it to walk on branches using its palms and were better suited for tool use. Thus, throughout further evolution, our ancestors did not need to significantly ‘remodel’ their hands.
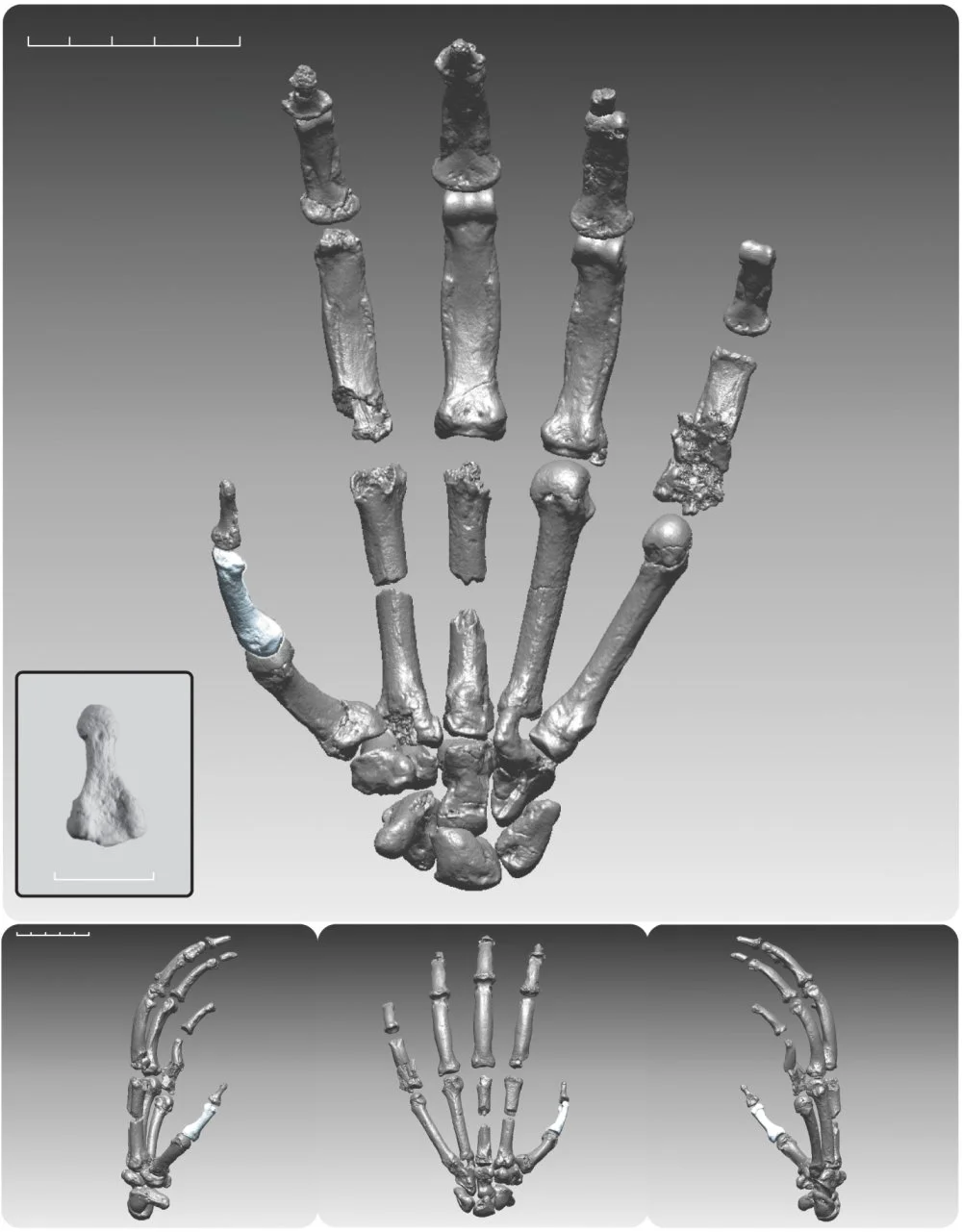
Digitally rendered composite hand of the “Ardi” partial skeleton. This image relates to an article in the 02 October 2009 issue of Science. The study, by Dr. C. Owen Lovejoy of Kent State University and colleagues, is titled, “Careful Climbing in the Miocene: The Forelimbs of Ardipithecus ramidus and Humans Are Primitive.” Image courtesy of Science/AAAS
The foot structure of Ardipithecus shows a mosaic of traits indicating both the retention of the ability to grasp branches (opposable big toe) and effective bipedalism (a stiffer arch than in modern great apes). Ardipithecus descendants, the australopithecines, lost the ability to grasp branches with their feet and developed a foot structure almost entirely like that of modern humans.
Ardipithecus also presented anthropologists with many other surprises. For example, it had never occurred to anyone that our ancestors initially adapted to bipedal walking through changes in the pelvis and only later lost the opposable big toe and grasping function of the feet.
Thus, the study of Ardipithecus revealed that some popular hypotheses about hominid evolution need revision.
Many traits of modern great apes turned out not to be primitive but advanced, including specific features of chimpanzees and gorillas related to deep specialization in climbing trees, hanging from branches, knuckle-walking, and specialized diets. These traits were not present in the common ancestors we share with them. The apes from which humans evolved were not very similar to present-day apes.
This likely pertains not only to physical structure but also to behavior and social organization. The thinking and social relations of chimpanzees may not be a good model for reconstructing the thinking and social relations of our ancestors. It seems incorrect to imagine our direct ancestors, the Australopithecines and Ardipithecus, as something like chimpanzees who learned how to walk upright. It appears that chimpanzees and gorillas are highly specialized, relict primates that have survived in impenetrable tropical forests, and this allowed them to persist to the present day. Based on new facts, American anthropologist Owen Lovejoy and his colleagues, who actively participated in the study of Ardipithecus, have developed a new model of early hominid evolution, which will be discussed in the next lecture. For now, it's important to highlight that love and tenderness played far more significant roles in the development of our species than one might assume, according to recent discoveries.